Reading time: 5 minutes
Keighley Reisenauer
Breast cancer is a collection of diseases, organized into subtypes designating which treatments may be best for the patient and how aggressive the cancer is likely to be. One thing that is universal across all subtypes, though, is what makes them worse: metastasis. When cancer metastasizes, it spreads across the body, hiding away in blood, lymph nodes, and other organs. Unfortunately, metastasis is the number one reason that cancer patients do not survive their disease; in breast cancer, it is cause for 90% of deaths1. Sometimes, those disseminated cells grow and divide quickly, forming macrometastases and large tumors that then themselves can start shedding cells into the body. Other times, those cells lie dormant, only to reawaken decades after the patient had achieved remission. To date, little is understood about the initial and long-term survival of these dormant breast cancer cells, making them even more difficult to treat and eliminate. Recently, though, a group from the NIH in Maryland has uncovered a non-intuitive driver for the survival of dormant breast cancer cells2.
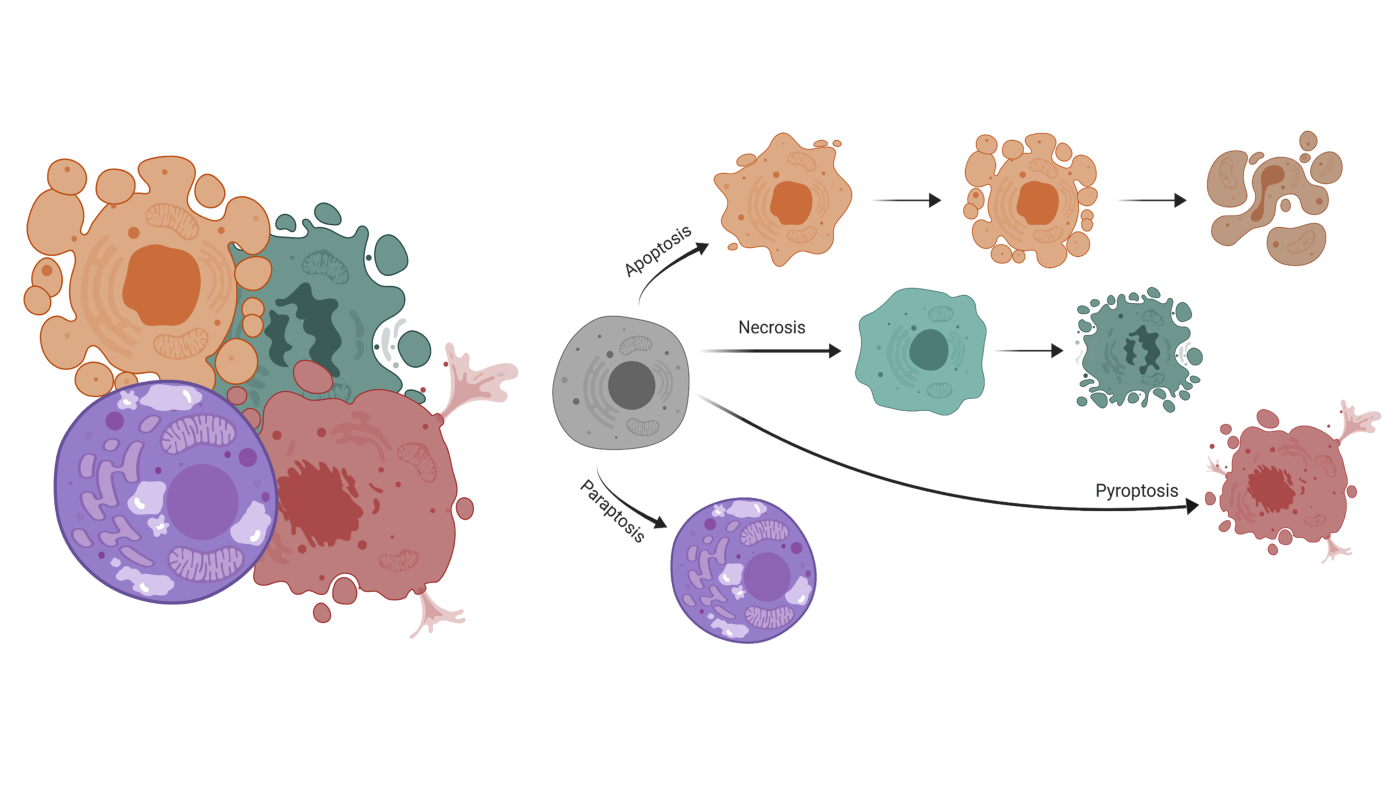
Many conventional chemotherapies and cancer treatments rely on activating mechanisms of cell death to destroy cancer cells. They rely on sensitivities in cancer cells that other healthy cells may be more robust against, such as deactivating key proteins, turning off important driver genes, or overloading the cell with signaling molecules. The end result is all roughly the same: the cell dies in a specific and coordinated manner. In the 2018 paper from Vera-Ramirez, et. al, authors describe the reliance on a cell death pathway in maintaining dormant cell survival in breast cancer. The most common type of cell death is apoptosis, by which cells receive a “death signal,” which originates when the cell is under stress and cannot rectify the situation. The molecules in the “death signal” start a process known as blebbing, where the cell starts to break apart in a manner that is not dissimilar to a lava lamp. The most common cellular stresses that initiate apoptosis are DNA damage, endoplasmic reticulum stress from misfolded protein accumulation, and cellular infection (ex: virus)3. Cells can also undergo necrosis, which is when the cell begins to swell and the plasma membrane breaks. Although there are different molecules involved, necrosis is also activated via a signaling pathway indicating irreparable damage to the cell, the presence of viral DNA in the cytosol, or some forms of DNA damage3. The type that the authors discover to be important for dormant cell survival, however, is autophagy. Autophagy (from the Greek “auto” meaning self, and “phagy”, to eat) is the least understood of the three major types of cell death, and it is usually activated in response to metabolic crisis or to remove damaged organelles3. Interestingly, autophagy activation results in the recycling of nutrients and energy, which could explain why this type of cell death is common in the interior spaces of tumors that are nutrient deprived. Cells self-sacrifice so that other nearby cells can continue to grow and divide, supplying the tumor with proliferative capacity4. In fact, multiple other studies have linked autophagy to additional tumorigenic properties such as metastasis5,6. For the first time, though, autophagy has been experimentally linked to the survival of dormant cells both in cell- and mouse-models of breast cancer2.
The NIH researchers used both cell- and mouse-models to experimentally determine the reliance on autophagy for dormant breast cancer cells. First, they demonstrated that their cell culture models were able to undergo autophagy in both permanently dormant and inducible proliferative backgrounds, where the measured autophagy signal decreased following the cells’ entry into a proliferative state. Next, they evaluated the effects of an autophagy inhibitor (Hydroxychloroquine, 50μM dose) on the viability of dormant cells. Following treatment with the inhibitor, regardless of the time point the treatment was applied, the dormant cells continued to die off, but the cells that were allowed to activate proliferation did not. The team was also able to demonstrate that application of the inhibitor in mice experiencing metastases was able to reduce outgrowth, even in an inducible proliferation cell model, suggesting that the autophagy inhibitor was able to prevent dormant cells from reawakening and metastasizing to the lung. Importantly, though, the treatment had to be applied quickly, before the cells were allowed to switch from dormant to proliferative. The paper rounds out the data by evaluating different gene expression in pathways associated with autophagy activation and dormancy. Data showed that autophagy was early to activate in the dormancy cell model, which was reversed when the inhibitor was applied.
Overall, authors describe the impact of inhibiting autophagy on the selective destruction of dormant cells but not proliferating cells. This underscores the importance and specificity of autophagic block in maintaining dormant cells in the non-proliferative state. Having disseminated tumor cells does not inherently increase a patient’s risk of disease recurrence. It is only when those cells activate and begin to grow out into tumors does the patient face a grim prognosis. The authors point out two interesting observations: (1) proliferative cells, either cancerous, normal, or reawakened dormant cells, are not responsive to autophagy inhibition and (2) cells undergo a quiescent state in the early stages of metastatic colonization of a secondary site during which the cells highly depend on autophagy. These caveats may explain previous discordant results in clinical trials evaluating autophagy and dormancy. The clarifying results produced by Vera-Ramirez, et. al. ultimately shed light on a potential mechanism for preventing the activation of dormant cells, thereby extending the remission status of breast cancer survivors.
Edited by Kayla Snare
Primary Work Discussed
Vera-Ramirez, L., Vodnala, S. K., Nini, R., Hunter, K. W. & Green, J. E. Autophagy promotes the survival of dormant breast cancer cells and metastatic tumour recurrence. Nature Communications, 9, 1944 (2018).
References
- Barkan, D. et al. Inhibition of metastatic outgrowth from single dormant tumor cells by targeting the cytoskeleton. Cancer Res. 68, 6241–6250 (2008).
- Vera-Ramirez, L., Vodnala, S. K., Nini, R., Hunter, K. W. & Green, J. E. Autophagy promotes the survival of dormant breast cancer cells and metastatic tumour recurrence. Nat Commun 9, 1944 (2018).
- Green, D. R. & Llambi, F. Cell Death Signaling. Cold Spring Harb Perspect Biol 7, a006080 (2015).
- Liu, J. & Debnath, J. The Evolving, Multifaceted Roles of Autophagy in Cancer. Adv. Cancer Res. 130, 1–53 (2016).
- Mowers, E. E., Sharifi, M. N. & Macleod, K. F. Autophagy in cancer metastasis. Oncogene 36, 1619–1630 (2017).
- White, E. The role for autophagy in cancer. J Clin Invest 125, 42–46 (2015).
Other Resources/Further Reading:
- Blank, M., and Shiloh, Y. (2007). Programs for Cell Death: Apoptosis is Only One Way to Go. Cell Cycle 6, 686–695.
- de Bruin, E.C., and Medema, J.P. (2008). Apoptosis and non-apoptotic deaths in cancer development and treatment response. Cancer Treatment Reviews 34, 737–749.
- Castro-Obregón, S., del Rio, G., Chen, S.F., Swanson, R.A., Frankowski, H., Rao, R.V., Stoka, V., Vesce, S., Nicholls, D.G., and Bredesen, D.E. (2002). A ligand-receptor pair that triggers a non-apoptotic form of programmed cell death. Cell Death & Differentiation 9, 807–817.
- Guaman-Ortiz, L., Orellana, M., and Ratovitski, E. (2017). Natural Compounds As Modulators of Non-apoptotic Cell Death in Cancer Cells. Current Genomics 18, 132–155.
- Lee, D., Kim, I.Y., Saha, S., and Choi, K.S. (2016). Paraptosis in the anti-cancer arsenal of natural products. Pharmacology & Therapeutics 162, 120–133.
- Mathiasen, I.S., and Jäättelä, M. (2002). Triggering caspase-independent cell death to combat cancer. Trends in Molecular Medicine 8, 212–220.
- Singha, P.K., Pandeswara, S., Venkatachalam, M.A., and Saikumar, P. (2013). Manumycin A inhibits triple-negative breast cancer growth through LC3-mediated cytoplasmic vacuolation death. Cell Death & Disease 4, e457–e457.
- Sperandio, S., de Belle, I., and Bredesen, D.E. (2000). An alternative, nonapoptotic form of programmed cell death. Proceedings of the National Academy of Sciences 97, 14376–14381.
- Sperandio, S., Poksay, K., de Belle, I., Lafuente, M.J., Liu, B., Nasir, J., and Bredesen, D.E. (2004). Paraptosis: mediation by MAP kinases and inhibition by AIP-1/Alix. Cell Death & Differentiation 11, 1066–1075.
- Tait, S.W.G., Ichim, G., and Green, D.R. (2014). Die another way – non-apoptotic mechanisms of cell death. Journal of Cell Science 127, 2135–2144.
- Ubah, O., and Wallace, H. (2014). Cancer Therapy: Targeting Mitochondria and other Sub-cellular Organelles. Current Pharmaceutical Design 20, 201–222.
- Yoon, M.J., Kim, E.H., Lim, J.H., Kwon, T.K., and Choi, K.S. (2010). Superoxide anion and proteasomal dysfunction contribute to curcumin-induced paraptosis of malignant breast cancer cells. Free Radical Biology and Medicine 48, 713–726.
- J. Kale, E.J. Osterlund, D.W. Andrews, BCL-2 family proteins: changing partners in the dance towards death, Cell Death Differ. 25 (2018) 65–80, https://doi.org/10.1038/cdd.2017.186.

Leave a comment